Centrala dogmen förklarad: DNA → RNA → protein i molekylärbiologin
Centrala dogmen: hur DNA → RNA → protein fungerar. Enkel, tydlig förklaring av replikation, transkription och translation för studenter och forskare.
Den centrala dogmen inom molekylärbiologin är ett begrepp myntat av Francis Crick, en av dem som föreslog DNA:s dubbelhelixstruktur. Den beskriver hur den genetiska informationen normalt flödar i celler: från DNA till RNA och vidare till protein. Crick formulerade idén första gången 1958 och preciserade den ytterligare 1970.
Begreppet är i grunden en ram för att förstå överföring av sekvensinformation mellan de tre huvudtyperna av biopolymerer i cellen: DNA, RNA (båda nukleinsyror) och protein. Utifrån dessa tre finns 3×3 = 9 teoretiskt möjliga direkta informationsöverföringar (från varje typ till varje annan). Crick delade in dem i tre grupper om tre:
Bildgalleri
5 Bilder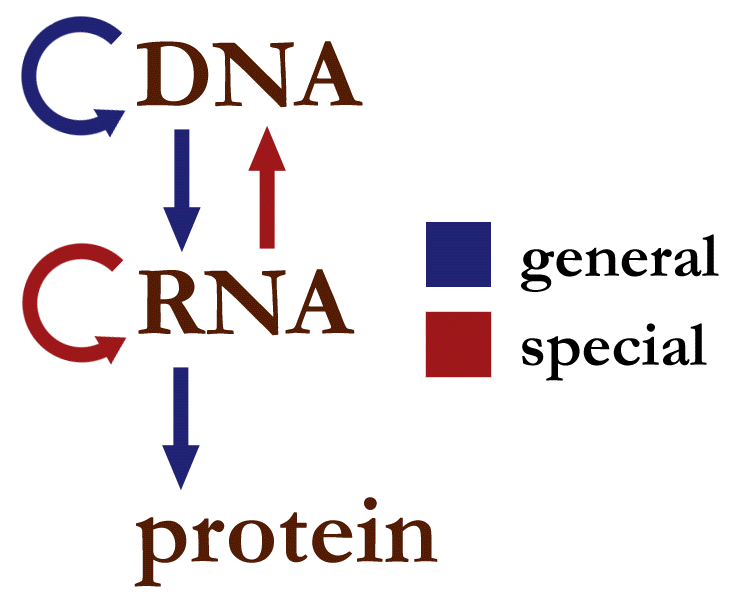



De tre grupperna av informationsöverföringar
- 3 allmänna överföringar – sker normalt i de flesta celler:
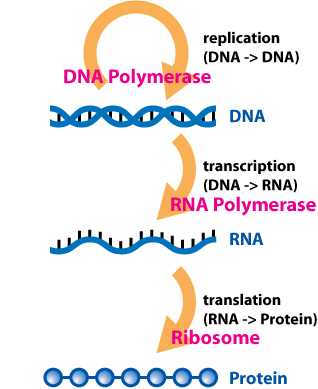
- DNA → DNA: DNA-replikation, kopiering av DNA vid celldelning.
- DNA → RNA: Transkription, då DNA-sekvensen avläses och kopieras till mRNA.
- RNA → protein: Översättning, ribosomerna använder mRNA som mall för proteinsyntes.
- 3 särskilda överföringar – kända att förekomma, men endast under speciella omständigheter (t.ex. hos vissa virus eller i laboratoriet):
- DNA → protein: direkt översättning från DNA utan mRNA-mellanprodukt. Detta är teoretiskt tänkbart men saknar goda naturliga exempel; normalt krävs RNA-mall.
- RNA → DNA: Reverse transcription, exempelvis hos retrovirus där enzymet revers transkriptas gör RNA till DNA.
- RNA → RNA: RNA-replikation som sker i många RNA-virus (RNA-beroende RNA-polymeras).
- 3 okända/omedgivet överföringar (enligt ursprunglig definition) – Crick ansåg att dessa i praktiken inte borde ske:
- Protein → protein: överföring av sekvensinformation från protein till protein. Modern forskning visar att proteiner kan överföra konformationell information (se prioner), men detta är inte en överföring av nukleotidsekvens.
- Protein → RNA: ingen välbelagd mekanism finns för att ett protein direkt skulle templatera en RNA-sekvens.
- Protein → DNA: inga kända exempel på ett protein som direkt kodar en nukleotidsekvens tillbaka till DNA.
Viktiga undantag och nyare insikter
Den centrala dogmen har ibland missförståtts som ett absolut förbud mot alla interaktioner mellan protein och nukleinsyror. Ett viktigt förtydligande är att dogmen gäller sekvensinformation — proteiner kan påverka genetiskt material på många sätt (till exempel som enzymer som modifierar DNA eller genom reglering av genuttryck), men detta är inte samma sak som att ett proteins aminosyrasekvens direkt överförs som nukleotidsekvens tillbaka till DNA eller RNA.
Några konkreta undantag och tillägg som upptäckts efter Cricks formulering:
- Reverse transcription (RNA → DNA): väl dokumenterat hos retrovirus och i laboratoriet (t.ex. med revers transkriptas).
- RNA → RNA replikation: vanligt hos många RNA-virus.
- Protein → protein (prioner): prioner visar att ett protein kan överföra sin tredimensionella konformation till ett annat protein och därigenom förändra dess funktion — detta är en form av informationöverföring men inte en överföring av nukleotidsekvens.
- Epigenetiska och kemiska modifieringar: proteiner kan förändra DNA:s metylering eller kromatinstruktur, vilket påverkar uttryck utan att ändra sekvensen; detta är en informationspåverkan men inte en sekvensöverföring från protein till nukleinsyra.
Varför dogmen är viktig
Den centrala dogmen ger en enkel och kraftfull modell för hur genetisk information används och hur livets molekylära maskineri är organiserat. Den är en grundsten i modern genetik, molekylärbiologi och bioteknik — bland annat för förståelse av sjukdomar orsakade av virus (t.ex. retrovirus), för utveckling av diagnostik och för metoder som reverse transcription–PCR.
Samtidigt är det viktigt att förstå både begränsningarna och nyanserna: dogmen beskriver huvuddelen av informationsflödet, men biologin innehåller undantag och ytterligare mekanismer som forskningen fortfarande utforskar.
Dogmen förknippas också med en äldre idé om informationsbarriärer i biologin, liknande August Weismanns princip (Weismann-barriären) — att ärftlig information normalt förs från könsceller till somatiska celler och inte i motsatt riktning. Detta perspektiv ligger fortfarande till grund för mycket av vår förståelse av arv och utveckling, även om fältet epigenetik visat att det finns fler lager av reglering än man trodde tidigare.

Frågor och svar
F: Vad är den centrala dogmen inom molekylärbiologin?
S: Molekylärbiologins centrala dogm är en fras av Francis Crick som säger att information överförs från DNA till proteiner via RNA, men att proteiner inte kan överföra informationen tillbaka till DNA.
F: När skrevs den för första gången?
S: Den centrala dogmen skrevs första gången av Francis Crick 1958 och upprepades 1970.
F: Vad ger dogmen en ram för att förstå?
S: Den centrala dogmen ger en ram för att förstå överföringen av sekvensinformation mellan biopolymerer som DNA, RNA och protein.
Fråga: Hur många direkta informationsöverföringar kan ske mellan dessa biopolymerer?
Svar: Det finns 3×3 = 9 tänkbara direkta informationsöverföringar som kan ske mellan dessa biopolymerer.
Fråga: Vilka är de tre grupper som klassificerar dessa överföringar?
S: Dessa överföringar klassificeras i tre grupper - allmänna överföringar (tros ske normalt i de flesta celler), särskilda överföringar (kända för att ske, men endast under särskilda förhållanden när det gäller vissa virus eller i ett laboratorium) och okända överföringar (tros aldrig ske).
F: Vad beskriver de allmänna överföringarna?
S: De allmänna överföringarna beskriver det normala flödet av biologisk information - DNA kan kopieras till DNA (DNA-replikation), DNA-information kan kopieras till mRNA (transkription) och proteiner kan syntetiseras med informationen i mRNA som mall (översättning).
F: Vad är Weismannbarriären?
S: Weismannbarriären är en princip som föreslogs av August Weismann och enligt vilken ärftlig information endast rör sig från gener till kroppsceller och aldrig tvärtom. Ärftlig information rör sig endast från könsceller till somatiska celler.
Relaterade artiklar
Författare
AlegsaOnline.com Centrala dogmen förklarad: DNA → RNA → protein i molekylärbiologin Leandro Alegsa
URL: https://sv.alegsaonline.com/art/18034
Källor
- profiles.nlm.nih.gov : On protein synthesis.
- nature.com : "Central dogma of molecular biology"
- doi.org : 10.1038/227561a0
- pubmed.ncbi.nlm.nih.gov : 4913914
- esp.org : esp.org/books/weismann/germ-plasm/facsimile/