Alternativ splitsning (splicing) – definition, funktion och betydelse
Alternativ splitsning: definition, mekanismer, funktion och betydelse för proteinmångfald, sjukdomar och cancer — hur en gen kan ge upphov till flera proteiner.
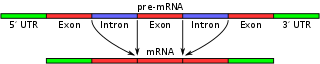
Vad är alternativ splicing?
Genom alternativ splicing kan en och samma gen ge upphov till flera olika proteiner. Detta sker genom att olika kombinationer av exoner inkluderas i det slutliga messenger RNA (mRNA) som bildas efter transkription.
Bildgalleri
10 Bilder
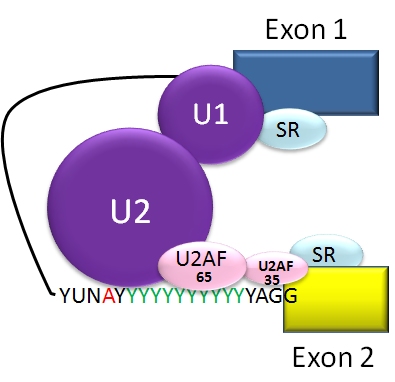
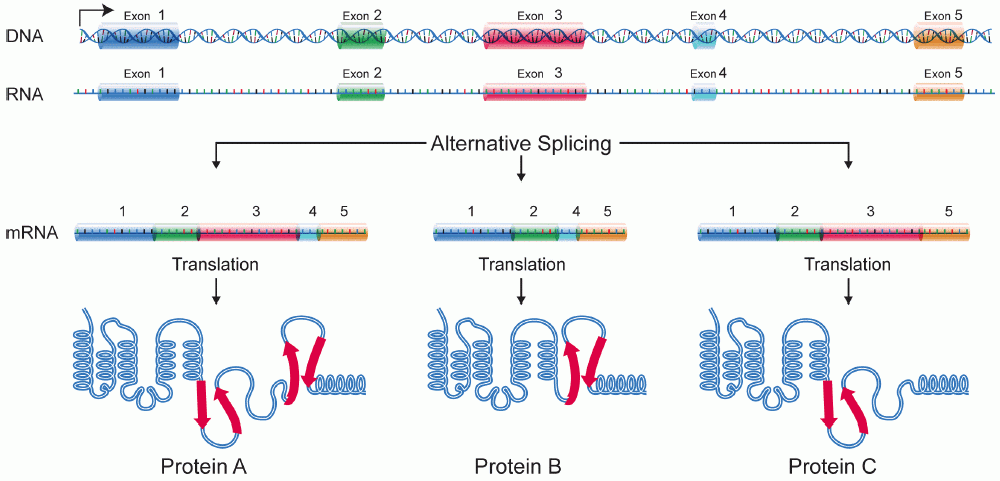





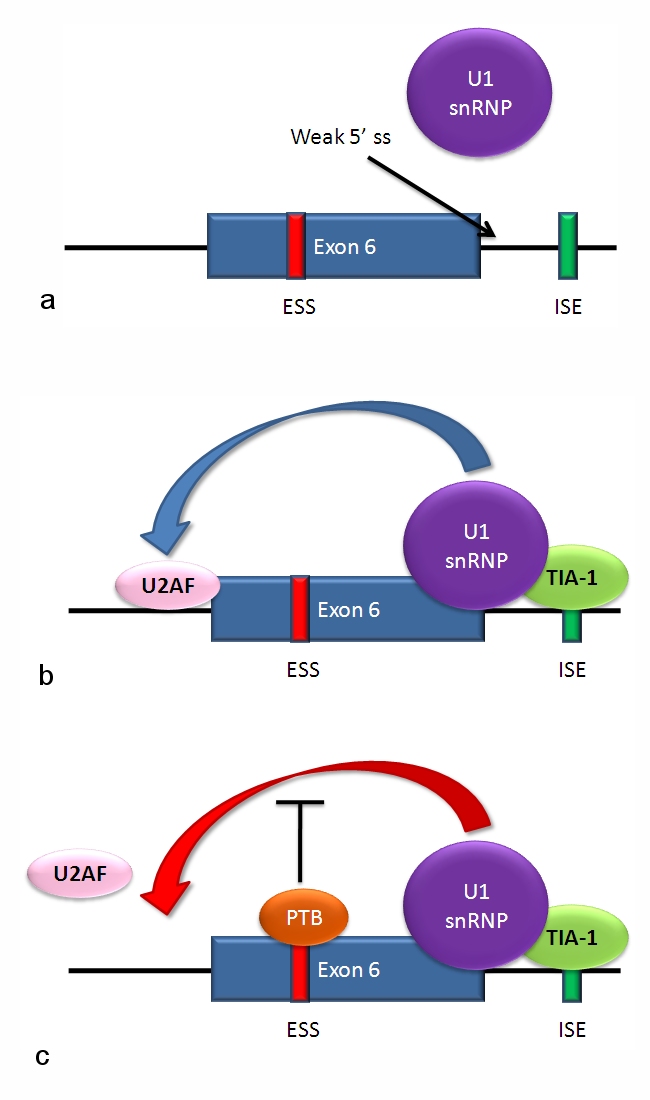
Hur går splicingen till?
Vid alternativ splicing kopplas exonerna i det pre‑mRNA som produceras genom transkription ihop på olika sätt under RNA‑splicing. Själva klippningen och hopsättningen utförs av spliceosomen, ett stort ribonukleoproteinkomplex bestående av små nukleära ribonukleoproteiner (snRNPs) och många andra proteiner. Spliceosomen känner igen konsensussekvenser vid exon‑intron‑gränserna (t.ex. 5'‑donor GU och 3'‑acceptor AG) samt en branch‑point och en polypyrimidin‑sträcka.
Typer av alternativ splicing
- Exonskippning (cassette exon): En exon kan inkluderas i vissa mRNA‑transkript men hoppas över i andra.
- Mutuellt exklusiva exoner: Endast en av två (eller flera) alternativa exoner ingår i varje transkript.
- Alternativt 5' splice‑site: Olika startpunkter för splicing vid exonets 5'‑ända ger kortare eller längre exon.
- Alternativt 3' splice‑site: Liknande, men med olika 3'‑ände.
- Intronretention: Ett intron behålls i det mogna mRNA:t, något som kan påverka läsram eller leda till nedbrytning.
Reglering av splicing
Valet av splicingmönster påverkas av både cis‑element i RNA och trans‑faktorer:
- Splicingaktivatorer och -repressorer: Korta sekvenser i pre‑mRNA, kallade exonskipping enhancers eller silencers, binder proteiner som stimulerar eller hindrar användningen av en viss spliceplats.
- Proteinfaktorer: SR‑proteiner (serin-/argininrika) fungerar ofta som aktivatorer, medan hnRNP‑proteiner ofta verkar som repressorer.
- Tidsmässig och vävnadsspecifik reglering: Många faktorer uttrycks olika i olika celltyper och utvecklingsstadier, vilket ger vävnadsspecifika splicingisoformer.
Funktionella konsekvenser
Alternativ splicing ökar proteomets komplexitet och kan påverka proteiners:
- funktion (olika enzymaktiviteter eller bindingsytor),
- subcellulära lokalisering (till exempel signalpeptid kan tas bort eller sättas in),
- stabilitet och nedbrytningsbenägenhet,
- interaktioner med andra proteiner och regleringsvägar.
På så sätt möjliggör alternativ splicing finjustering av cellens funktion och svar på miljö‑ eller utvecklingssignaler. Alternativ splicing är särskilt omfattande i eukaryoter och är en viktig orsak till att ett relativt begränsat antal gener kan ge upphov till ett mycket större antal proteiner. Hos människor är ungefär 95 % av de multi‑exoniska generna alternativt splicade.
Sjukdomar och klinisk betydelse
Onormala variationer i splicing förekommer vid sjukdom. Många genetiska sjukdomar har sin grund i felaktig splicing — mutationer kan förstöra en splice‑site, skapa en ny sådan eller förändra bindningsställen för regulatoriska proteiner. Onormala splicingvarianter kan också bidra till utvecklingen av cancer genom att skapa isoformer som främjar tillväxt eller överlevnad.
Det finns numera också terapier som riktar sig mot splicing:
- Antisense‑oligonukleotider (ASO): Kan användas för att blockera eller ändra ett splicingmönster (t.ex. exon‑skipping‑strategier för Duchenne muskeldystrofi).
- Splice‑switching läkemedel: Exempelvis nusinersen för spinal muskelatrofi (SMA), som modifierar splicing av SMN2‑transkript för att öka produktionen av funktionellt SMN‑protein.
- Små molekyler: Pågående utveckling av små molekylära läkemedel som påverkar spliceosomen eller splicingfaktorer.
Detektion och analys
Alternativ splicing studeras med flera metoder:
- RT‑PCR och kvantitativ PCR: Lämpligt för att påvisa och kvantifiera kända isoformer.
- RNA‑sekvensering (RNA‑seq): Ger en omfångsrik, genombred analys av splicingmönster och används för att upptäcka både kända och nya varianter.
- Bioinformatiska verktyg och databaser: Analyserar reads och förutsäger exon‑intron‑gränser och isoformer.
Kvalitetskontroll och nedbrytning av felaktiga transkript
Icke fungerande eller skadliga splicingprodukter hanteras vanligtvis genom cellens RNA‑kvalitetskontrollsystem. Ett viktigt sådant är nonsense‑mediated decay (NMD), som känner igen mRNA med prematura stoppkodon och dirigerar dem till nedbrytning. Andra system, som RNA‑exosomen och olika ribonukleaser, bidrar också till nedbrytning av skadade eller onödiga transkript — ofta genom att de "hackas upp" av olika enzymer.
Evolutionsperspektiv
Alternativ splicing bidrar till evolutionsmässig flexibilitet: genom att skapa nya kombinationer av domäner och funktionella moduler kan organismer snabbt få fram nya proteinvarianter utan att behöva nya gener. Splicingmönster kan vara konserverade eller snabbt föränderliga beroende på urvalstryck och funktionell betydelse.
Sammanfattning
Alternativ splicing är en central mekanism i eukaryoter som tillåter en och samma gen att producera flera olika mRNA‑isoformer och därmed flera proteiner. Genom komplex reglering av spliceosomen och bindande proteiner skapas vävnads‑ och situationsspecifika proteiner. Felaktig splicing är en viktig orsak till sjukdom, men samma mekanism erbjuder även terapeutiska möjligheter genom riktade behandlingar.

Källa till mångfald
Alternativ splicing (rekombination av olika exoner) är en viktig källa till genetisk mångfald hos eukaryoter. En viss Drosophila-gen (DSCAM) kan splicas alternativt till 38 000 olika mRNA.
Frågor och svar
F: Vad är alternativ splicing?
S: Alternativ splicing är en process där exonerna i pre-messenger RNA som produceras genom transkription återkopplas på olika sätt under RNA-splicing, vilket resulterar i olika mogna messenger RNA från samma gen som översätts till olika proteiner.
F: Hur vanligt är alternativ splicing?
S: Alternativ splicing är normalt hos eukaryoter och ökar kraftigt mångfalden av proteiner som kan kodas av genomet. Hos människor är omkring 95 % av de multiexoniska generna alternativt splicade.
F: Vilka är några typer av alternativ splicing?
S: Den vanligaste typen av alternativ splicing är exon skipping, där en exon kan inkluderas eller utelämnas från mRNA beroende på vissa förhållanden eller vävnader. Det finns också andra typer, t.ex. splicingaktivatorer och -repressorer som främjar respektive minskar användningen av vissa platser, samt nya typer som håller på att upptäckas.
F: Hur påverkar onormal variation i alternativ splicing hälsan?
S: Onormala variationer i alternativ splicing kan leda till genetiska störningar och bidra till cancerutveckling. Icke fungerande produkter som uppstår i denna process hanteras vanligtvis genom posttranskriptionell kvalitetskontroll, där de hackas upp av enzymer.
F: Vilken typ av proteiner kan skapas genom alternativ splicing?
S: Genom alternativ splicing kan en enda gen koda för flera proteiner, vilket ökar mångfalden av proteiner som kan kodas av genomet.
F: Vad händer om icke-arbetande produkter uppstår vid alternativ splejsning?
S: Om icke-arbetande produkter uppstår vid alternativ skarvning hanteras de vanligen genom posttranskriptionell kvalitetskontroll, där de hackas upp av enzymer.
Relaterade artiklar
Författare
AlegsaOnline.com Alternativ splitsning (splicing) – definition, funktion och betydelse Leandro Alegsa
URL: https://sv.alegsaonline.com/art/3063
Källor
- doi.org : 10.1146/annurev.biochem.72.121801.161720
- pubmed.ncbi.nlm.nih.gov : 12626338
- doi.org : 10.1038/ng.259
- pubmed.ncbi.nlm.nih.gov : 18978789
- doi.org : 10.1038/nrm1645
- pubmed.ncbi.nlm.nih.gov : 15956978
- doi.org : 10.1101/gad.1643108
- pubmed.ncbi.nlm.nih.gov : 18245441
- doi.org : 10.1016/j.biocel.2007.02.016
- pubmed.ncbi.nlm.nih.gov : 17416541
- ncbi.nlm.nih.gov : "A global view of cancer-specific transcript variants by subtractive transcriptome-wide analysis"
- doi.org : 10.1371/journal.pone.0004732
- pubmed.ncbi.nlm.nih.gov : 19266097
- ncbi.nlm.nih.gov : "Aberrant RNA splicing and its functional consequences in cancer cells"