Artbildning (speciering) – hur arter uppstår: definition, processer och exempel
Artbildning (speciering) – lär dig hur arter uppstår: definitioner, evolutionära processer, isolering, hybridisering och tydliga exempel för att förstå biologisk mångfald.
Artbildning handlar om hur arter bildas och skiljer sig från varandra. Det är en central fråga i evolutionsbiologin eftersom förståelsen av artbildning förklarar mångfalden av liv och hur nya biologiska former uppstår över tid.
I klassisk syn på evolution betonade Darwin gradvis förändring genom naturligt urval. Två begrepp som ofta används för att beskriva artutveckling är anagenesis och kladogenes. Anagenesis (ibland kallat fyletisk evolution) innebär att en hel population gradvis förändras så att den över tid blir så olika sina föregångare att den betraktas som en ny art. Kladogenes innebär istället att en art splittras i två eller flera linjer som sedan utvecklas självständigt — en förgrening i livets träd.
Bildgalleri
10 Bilder









Olika lägen för artbildning
Artbildning kan ske under olika geografiska och ekologiska förutsättningar. De vanligaste modellerna är:
- Allopatrisk artbildning – populationsdelning genom geografisk separation (t.ex. berg, hav eller avstånd). Isolering minskar eller upphäver genflöde, vilket tillåter genetisk differentiering genom mutation, naturligt urval och genetisk drift.
- Peripatrisk artbildning – en särskild form av allopatrisk artbildning där en liten delpopulation isoleras i utkanten av artens utbredning. Starka effekter av genetisk drift och snabb anpassning i en liten population kan påskynda artbildning.
- Parapatrisk artbildning – när grannpopulationer differentierar längs en miljögradient utan fullständig geografisk barriär; begränsat genflöde kan leda till differentiering.
- Sympatrisk artbildning – artbildning utan geografisk separation, ofta driven av ekologisk specialisering, sexuellt urval eller genetiska förändringar som polyploidi (särskilt vanlig hos växter).
Mekanismer bakom reproduktiv isolering
Att bli en ”ny art” innebär ofta att populationer uppnår någon grad av reproduktiv isolering. Denna isolering kan vara:
- Prezygotiska mekanismer – förhindrar befruktning (t.ex. olika parningstid, olika parningsbeteenden, mekaniska hinder eller gametisk inkompatibilitet).
- Postzygotiska mekanismer – påverkar avkommans överlevnad eller fertilitet efter befruktning (t.ex. sterila hybrider eller lägre hybridfitness).
Isolering kan uppstå gradvis genom naturligt urval (t.ex. förstärkning där selektion mot ofördelaktiga hybrider ökar prezygotisk isolering), eller genom slumpmässiga processer som genetisk drift, särskilt i små populationer.
Hybridisering, genflöde och modern genetik
Under de senaste decennierna har analyser av DNA-sekvensen hos levande organismer gett nya insikter. Många studier visar att hybridisering mellan närbesläktade arter är vanligare än man tidigare trodde. När arter korsar sig kan gener överföras mellan arter i en process som kallas introgression. Detta innebär att strikt reproduktiv isolering inte alltid är ett absolut kriterium för artgräns och att artbildning ibland sker trots fortsatt genflöde.
I praktiken kan hybridisering leda till:
- Introgression av adaptiva gener som underlättar snabb anpassning till nya miljöförhållanden.
- Hybridartbildning där korsningar mellan arter ger upphov till stabila, reproduktivt isolerade linjer (vanligt hos växter via polyploidi).
- Kollaps av artgränser när omfattande hybridisering suddar ut skillnaderna mellan tidigare separata arter.
Exempel och empiriska belägg
Flera välstuderade exempel visar olika aspekter av artbildning:
- Darwins finkar på Galápagos öar — klassiskt exempel på allopatrisk och adaptiv differentiering driven av olika födokällor och naturligt urval.
- Cichlider i Afrikas stora insjöar — extremt snabb artbildning och ekologisk specialisering, ofta med sympatriska eller parapatriska komponenter.
- Polyploid artbildning hos växter (t.ex. hos vissa vildgräs och blåklintsläkten) — nya arter kan uppstå över en generation genom heltaliga förändringar i kromosomantalet.
- Introgression hos människor — genetiska spår av korsningar med neandertalare och denisovaner visar att genflöde mellan distinkta befolkningslinjer kan lämna bestående arv.
- Ringarter och gradvisa förändringar längs utbredningsområden (t.ex. vissa salamandrar och måsfamiljer) illustrerar hur kontinuerliga förändringar kan leda till oförmåga att para sig i kontaktzonen.
Tidsramar och komplexitet
Tiden för artbildning varierar kraftigt: vissa arter kan skilja sig på relativt korta tidsskalor (tusentals till tiotusentals år eller snabbare i vissa fisk- och växtgrupper), medan andra linjer visar mycket långsam differentiering. Den biologiska verkligheten är ofta komplex: olika processer kan samverka och samma artgräns kan vara flytande beroende på vilken egenskap man studerar (morfologi, ekologi, genetik eller reproduktionsbarriärer).
Betydelse och nutida forskning
Studier av artbildning är viktiga för att förstå biodiversitet, bevarandebiologi och hur arter kan svara på miljöförändringar. Modern genetik och genomik har gjort det möjligt att spåra hur gener rör sig mellan populationer och arter, att identifiera gener som påverkar reproduktiv isolering och att upptäcka tidigare okända hybridiseringshändelser. Tillsammans visar dessa data att artbildning är en dynamisk process med många möjliga vägar, inte en enda linjär modell.
Sammanfattning: Artbildning kan ske genom gradvis förändring eller genom uppdelning i nya linjer. Geografisk separation (allopatri) är ofta viktig, men hybridisering, polyploidi, selektion och drift bidrar också. Moderna DNA-studier visar att gränserna mellan arter kan vara mer genomträngliga och komplexa än tidigare antaget.
Isoleringsmekanismer
Isoleringsmekanismer är saker som förhindrar framgångsrik avel mellan grupper inom en art. Reproduktiv isolering av populationer har etablerats. Detta är särskilt viktigt för det biologiska artbegreppet, eftersom arter definieras genom reproduktiv isolering.
Isoleringsmekanismerna kan delas in i två grupper, före och efter befruktningen.
Före gödsling
Faktorer som hindrar individer från att para sig.
- Geografisk isolering: Arter förekommer i olika områden och är ofta åtskilda av barriärer.
- Tidsmässig isolering: Individer parar sig inte eftersom de är aktiva vid olika tidpunkter. Det kan vara olika tider på dygnet eller olika årstider. Arternas parningsperioder kanske inte stämmer överens. Individer möter inte varandra under parningsperioderna, eller inte alls.
- Ekologisk isolering: Individer parar sig endast i den livsmiljö de föredrar. De möter inte individer av andra arter med andra ekologiska preferenser.
- Beteendemässig isolering: Individer av olika arter kan mötas, men man känner inte igen de sexuella signaler som kan ges. En individ väljer i de flesta fall en medlem av sin egen art.
- Mekanisk isolering: Kopulation kan försöka, men överföring av spermier sker inte. Individerna kan vara inkompatibla på grund av storlek eller morfologi.
- Spelteknisk inkompatibilitet: Spermaöverföring sker, men ägget befruktas inte.
Efter gödsling
Faktorer som förhindrar en lyckad parning, t.ex. genetisk inkompatibilitet, hybridavvikelse eller sterilitet.
- Zygotisk dödlighet: Ägget befruktas, men zygoten utvecklas inte.
- Hybrid som inte är livsduglig: Hybridembryot bildas, men är inte livsdugligt.
- Hybrid sterilitet: Hybrid är livskraftig, men den vuxna arten är steril.
- Hybridfördelning: Första generationens hybrider (F1) är livskraftiga och fertila, men ytterligare hybridgenerationer (F2 och backcross) är odugliga eller sterila.

Geografisk isolering
Detta anses vara den vanligaste orsaken till artbildning. Den första som kom på det var Moritz Wagner, en tysk upptäcktsresande och naturhistoriker.
Wagner började sin karriär som geograf och publicerade ett antal geografiska böcker om Nordafrika, Mellanöstern och Tropiska Amerika. Han var också en ivrig naturforskare och samlare, och det är för detta arbete han är mest känd bland biologer. Ernst Mayr, evolutionisten och biologihistorikern, har gett en redogörelse för Wagners betydelse.p562–565
Under sina tre år i Algeriet studerade Wagner (bland annat) de flyglösa skalbaggarna Pimelia och Melasoma. Varje släkte är uppdelat i ett antal arter som var och en är begränsad till en del av nordkusten mellan floderna som går ner från Atlasbergen till Medelhavet. Så snart man korsar en flod dyker en annan men närbesläktad art upp.
"... en begynnande art kommer endast att [uppstå] när några få individer överskrider gränserna för sitt utbredningsområde ... bildandet av en ny ras kommer aldrig att lyckas ... utan att kolonisterna under lång tid har separerats från de andra medlemmarna av sin art".
Detta var en tidig beskrivning av en process av en typ av geografisk artbildning. År 1942 återinfördes den av Mayr, och betydelsen av geografisk artbildning blev en av kärnidéerna i den evolutionära syntesen.
En annan term för geografisk artbildning är allopatrisk artbildning. Allopatry betyder "olika land".
Lediga öar
Vulkaniska öar bildas utan liv, och allt liv måste komma med hjälp av vind eller vatten. Vi vet från Hawaiiöarna och Galapagosöarna att alla former av liv förändras när de når öarna från fastlandet.
På Hawaiiöarna, som ligger på cirka 17 000 km2 , finns världens mest varierande samling av drosophilidflugor, som lever i allt från regnskogar till bergsängar. Det finns omkring 800 kända arter av drosophilider på Hawaii.
Studier visar att det finns ett tydligt "flöde" av arter från äldre till nyare öar. Det finns också fall av kolonisering tillbaka till äldre öar och överhoppning av öar, men dessa är mycket ovanligare.
Med hjälp av kalium/argon-radioaktiv datering kan de nuvarande öarna dateras från 0,4 miljoner år sedan (mya) (Mauna Kea) till 10 miljoner år sedan (Necker). Den äldsta delen av Hawaiis arkipelag som fortfarande befinner sig ovanför havet är Kure Atoll, som kan dateras till 30 mya.
Själva skärgården, som uppstod när Stillahavsplattan rörde sig över en het punkt, har funnits mycket längre, åtminstone under kritaperioden. Hawaii-öarna plus tidigare öar som nu ligger under havet utgör Hawaii- och kejsarbergskedjan, och många av undervattensbergen är guyots.
Alla inhemska drosophilid-arter på Hawaiʻi härstammar tydligen från en enda stamart som koloniserade öarna för cirka 20 miljoner år sedan. Den efterföljande adaptiva strålningen sporrades av bristen på konkurrens och ett brett utbud av lediga nischer. Även om det skulle vara möjligt för en enskild dräktig hona att kolonisera en ö, är det mer troligt att det har varit en grupp från samma art.
Det finns andra djur och växter i Hawaiis arkipelag som har genomgått liknande, om än mindre spektakulära, anpassningsstrålningar.
Ringarter
Inom biologin är en ringart en sammanhängande serie av närliggande populationer som var och en kan korsa sig med närliggande populationer. De två ändarna av kedjan överlappar varandra.
De två slutpopulationerna i serien är för långt ifrån varandra för att kunna korsas. Sådana "slutpopulationer" som inte är avelsförenade, men som är genetiskt förbundna, kan samexistera i samma region och på så sätt sluta en "ring".
Ringarter är viktiga bevis på evolution: de visar vad som händer med tiden när populationer genetiskt skiljer sig åt. Richard Dawkins påpekade att ringarter "endast visar oss i den rumsliga dimensionen något som alltid måste ske i tidsdimensionen".
Det är dock svårt att hitta ett enkelt och okomplicerat exempel.
Larus måsar
Detta var ett klassiskt exempel på ringarter. Dessa måsars utbredningsområde bildar nästan en ring runt Nordpolen (som normalt inte överflygs av måsar). Den mindre svartryggade måsen och fiskmåsarna är tillräckligt olika för att de normalt inte hybridiserar; så, sa man, gruppen av måsar bildar ett kontinuum utom där de två linjerna möts i Europa. Man är dock allmänt överens om att detta inte är helt korrekt, även om detaljerna är ytterst komplicerade.
Ensatina salamandrar
Ensatina-salamandern är en ringart i bergen runt Kaliforniens centrala dalgång. Komplexet bildar en hästskoform runt bergen. Även om korsning kan ske mellan var och en av de 19 populationerna runt hästskon, kan underarten Ensatina eschscholtzii i den västra änden av hästskon inte korsa sig med Ensatina klauberi i den östra änden. Det är en illustration av "nästan alla stadier i en artbildningsprocess" (Dobzhansky). Richard Highton hävdade att Ensatina är ett fall av flera arter och inte ett kontinuum av en art.
Den grönaktiga sångerskan
Grönsångaren (Phylloscopus trochiloides) har ett antal underarter, av vilka P. t . viridianus är den mest kända i Europa. Det är en ringart med populationer som divergerar öster och väster om den tibetanska högplatån för att senare mötas på den norra sidan. Deras relationer är ganska förvirrande.




Sympatrisk artbildning
Sympatrisk artbildning innebär att två eller flera arter bildas från en enda stamart som alla lever på samma geografiska plats. Detta anses nu vara mycket vanligt.
Vid sympatrisk artbildning skiljer sig arterna åt medan de bor på samma plats. Ofta citerade exempel på sympatrisk artbildning finns hos insekter som blir beroende av olika värdväxter i samma område.
Existensen av sympatrisk artbildning som en mekanism för artbildning var starkt ifrågasatt. Man hävdade att bevisen för sympatrisk artbildning i själva verket är exempel på mikrogeografisk artbildning. I allmänhet är detta nu inte den föredragna förklaringen. Det är snarare resultatet av hybridisering mellan närbesläktade arter, följt av naturligt urval som verkar på avkomman från sådana korsningar. Ett allmänt accepterat exempel på sympatrisk artbildning är cichliderna i Nabugabosjön i Östafrika, som tros bero på sexuellt urval.
Artbildning genom polyploidisering
Polyploiditet har orsakat många snabba artbildningar eftersom avkommor från t.ex. tetraploida x diploida parningar ofta resulterar i triploida sterila avkommor.
Alla polyploider är dock inte reproduktivt isolerade från sina föräldraväxter, och genflöde kan fortfarande förekomma, till exempel genom triploida hybrid x diploida parningar som ger upphov till tetraploider.
Många av de existerande växt- och de flesta djurarter har uppenbarligen genomgått polyploidisering under sin evolutionära historia. Reproduktionen av framgångsrika polyploida arter är ibland asexuell, genom parthenogenes. Av okända skäl är många asexuella organismer polyploida.
Hagtornsfluga
Ett exempel på hur evolutionen fungerar är fallet med hagtornsflugan Rhagoletis pomonella, som verkar genomgå sympatrisk artbildning.
Olika populationer av hagtornsflugor äter olika frukter. En särskild population uppstod i Nordamerika på 1800-talet, en tid efter att äpplen, som är en främmande art, introducerades. Denna äppelätande population livnär sig normalt sett endast på äpplen och inte på den historiskt sett föredragna frukten hagtorn. Den nuvarande populationen som äter hagtorn äter normalt inte äpplen.
Vissa bevis tyder på att sympatrisk artbildning förekommer. Hagtornsflugor mognar senare under säsongen och tar längre tid på sig än äppelflugor, och det finns få bevis för korsning (forskare har dokumenterat en hybridiseringsgrad på 4-6 %).
Uppkomsten av den nya hagtornsflugan är ett exempel på pågående evolution.
Hybridisering
I sällsynta fall bildas en ny art när enskilda individer av olika arter parar sig. Vanligtvis är produkterna av sådana hybridparningar infertila (inte fertila), eller relativt infertila, och elimineras därför av det naturliga urvalet.
Ett exempel på en ny och framgångsrik hybridart har hittats. Den nya arten är en hybrid mellan den italienska och den spanska sparven, och de grundläggande fakta har kontrollerats genom sekvensanalys av DNA:t i deras blod. Fågeln lever i Italien i en region där båda föräldraarterna lever. Den förökar sig inte med den spanska sparven trots att den lever sida vid sida med dem.
Artificiell artbildning
Nya arter har skapats genom domesticerad djurhållning, men de första tidpunkterna och metoderna är oklara. Tamfåren skapades till exempel genom hybridisering och ger inte längre livskraftig avkomma med Ovis orientalis, en art som de härstammar från.
Däremot kan tamdjur betraktas som samma art som flera olika sorter av vilda oxar, gaur, jakar etc., eftersom de lätt producerar fertila avkommor tillsammans med dem.
Labbetsarter
De bäst dokumenterade skapelserna av nya arter i laboratoriet gjordes i slutet av 1980-talet. William Rice och G.W. Salt avlade fruktflugor, Drosophila melanogaster, med hjälp av en labyrint med tre olika val av livsmiljöer, t.ex. ljus/mörker och vått/torrt.
Varje generation placerades i labyrinten och de grupper av flugor som kom ut genom två av de åtta utgångarna sattes åtskilda för att föröka sig med varandra i sina respektive grupper. Efter trettiofem generationer var de två grupperna och deras avkomma reproduktivt isolerade på grund av deras starka habitatpreferenser: de parade sig endast inom de områden de föredrog, och parade sig därför inte med flugor som föredrog de andra områdena. Historien om sådana försök beskrivs i Rice och Hostert (1993).
Diane Dodd kunde också visa hur reproduktiv isolering kan utvecklas från parningspreferenser hos Drosophila pseudoobscura efter bara åtta generationer med hjälp av olika typer av föda, stärkelse och maltos.
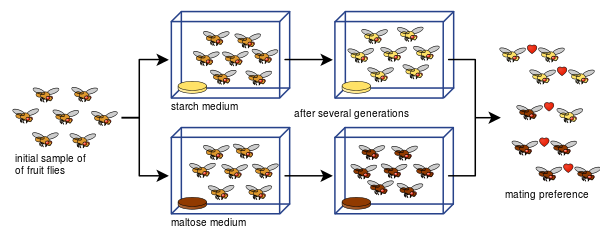
Det har varit lätt för många andra att kopiera Dodds experiment, även med andra sorters fruktflugor och livsmedel.
Förstärkning (Wallace-effekt)
Förstärkning är den process genom vilken det naturliga urvalet ökar den reproduktiva isoleringen.
Den kan uppstå när två populationer av samma art separeras och sedan återigen kommer i kontakt med varandra. Om deras reproduktiva isolering var fullständig har de redan utvecklats till två separata inkompatibla arter.
Om deras reproduktiva isolering är ofullständig kommer ytterligare parning mellan populationerna att ge upphov till hybrider, som kan vara fertila eller inte. Om hybriderna är infertila, eller fertila men mindre vältränade än sina förfäder, kommer det inte att ske någon ytterligare reproduktiv isolering och artbildning har i huvudsak ägt rum (t.ex. som hos hästar och åsnor).
Resonemanget bakom detta är att om föräldrarna till hybridavkomman var och en har naturligt utvalda egenskaper för sina egna olika miljöer, kommer hybridavkomman att bära egenskaper från båda och inte passa in i någon av nischerna lika bra som någon av föräldrarna. Den låga lämpligheten hos hybriderna skulle leda till att urvalet gynnar assortativ parning, vilket skulle minska hybridiseringen.
Detta kallas ibland för Wallace-effekten, efter evolutionsbiologen Alfred Russel Wallace som i slutet av 1800-talet föreslog att det kunde vara en viktig faktor för artbildning.
Om hybridavkommorna är mer välmående än sina förfäder kommer populationerna att smälta samman till samma art inom det område där de är i kontakt med varandra.
Förstärkning krävs för separation när det finns en "hybridzon" mellan två former av en art. Hybridzoner är områden där skilda populationer möts och korsar varandra. Hybridavkommor är mycket vanliga i dessa områden, som vanligtvis skapas genom att skilda arter kommer i sekundär kontakt.
Utan förstärkning skulle de två populationerna eller arterna ha okontrollerbar korsning. Förstärkning kan framkallas i artificiella urvalsexperiment enligt beskrivningen ovan.
Frågor och svar
F: Vad är artbildning?
S: Speciation är processen för hur arter bildas. Det är en viktig del av evolutionsbiologin.
F: Vad tyckte Darwin om artbildning?
S: Darwin trodde att de flesta arter kom direkt från redan existerande arter, vilket han kallade anagenesis eller "phyletisk evolution".
F: Hur såg man allmänt på artbildning på 1900-talet?
S: På 1900-talet trodde forskarna att de flesta arter bildades när tidigare arter splittrades, vilket kallades för kladogenes. Man trodde också att denna uppdelning orsakades eller underlättades av isoleringsmekanismer som fysisk separation.
F: Hur har den senaste forskningen förändrat vår förståelse av artbildning?
S: Ny forskning har visat att hybridisering mellan besläktade arter kan förekomma och att gener kan överföras genom dessa korsningar. Detta innebär att reproduktiv isolering inte alltid är nödvändig för att definiera en art och att allopatry (fysisk separation) kanske inte är nödvändigt för att artbildning ska äga rum.
F: Vad har fysisk separation med bildandet av nya arter att göra?
S: Fysisk separation mellan arter som en gång existerade tillsammans tros vara en viktig faktor för bildandet av nya arter, vilket illustreras av många exempel som diskuteras i den vetenskapliga litteraturen.
Relaterade artiklar
Författare
AlegsaOnline.com Artbildning (speciering) – hur arter uppstår: definition, processer och exempel Leandro Alegsa
URL: https://sv.alegsaonline.com/art/92570
Källor
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833